Open your refrigerator. Somewhere on a shelf — between the milk and the leftovers, shrink-wrapped and unremarkable — there is a dinosaur.
Not a descendant of one. Not a distant relative with a flattering family story. A dinosaur, in the same precise sense that you are a mammal: a member of the group, by descent, with no opt-out clause. The chicken is a theropod — the same branch of the dinosaur tree that produced Tyrannosaurus and Velociraptor — and it has been sitting in the most boring corner of your kitchen the entire time.

I know how this sounds. It sounds like the kind of "well, actually" that a clever teenager deploys to feel superior — a word game dressed up as a discovery. So let me promise you two things up front. First, the strongest version of that objection is real, and we are going to give it a whole section and answer it honestly, not wave it away. Second, by the end, the thing that will surprise you is not that the claim survives scrutiny — it's how much evidence had to pile up before scientists were forced to accept something this strange. The chicken isn't a dinosaur because somebody decided to call it one. It's a dinosaur because every attempt to rule that out eventually fell apart.
Part 1"Is," not "descended from"
Here's the move that trips everyone, including people who like dinosaurs: birds evolved from dinosaurs feels humble and correct, and birds are dinosaurs feels like an overreach. Surely the dinosaurs are the dead ones, and the birds are what came after.
But that's not how ancestry works once you follow it strictly. You don't graduate out of the family you were born into. A bat evolved a radically new body — powered flight, echolocation, the works — and a bat is still, fully and permanently, a mammal. A whale went back to the sea and turned its legs into flippers, and a whale is still an artiodactyl, the same group as deer and hippos. Descendants keep their inheritance. There is no act of becoming-so-different that ejects a lineage from its ancestral group; the group is defined as "the common ancestor and all of its descendants."1
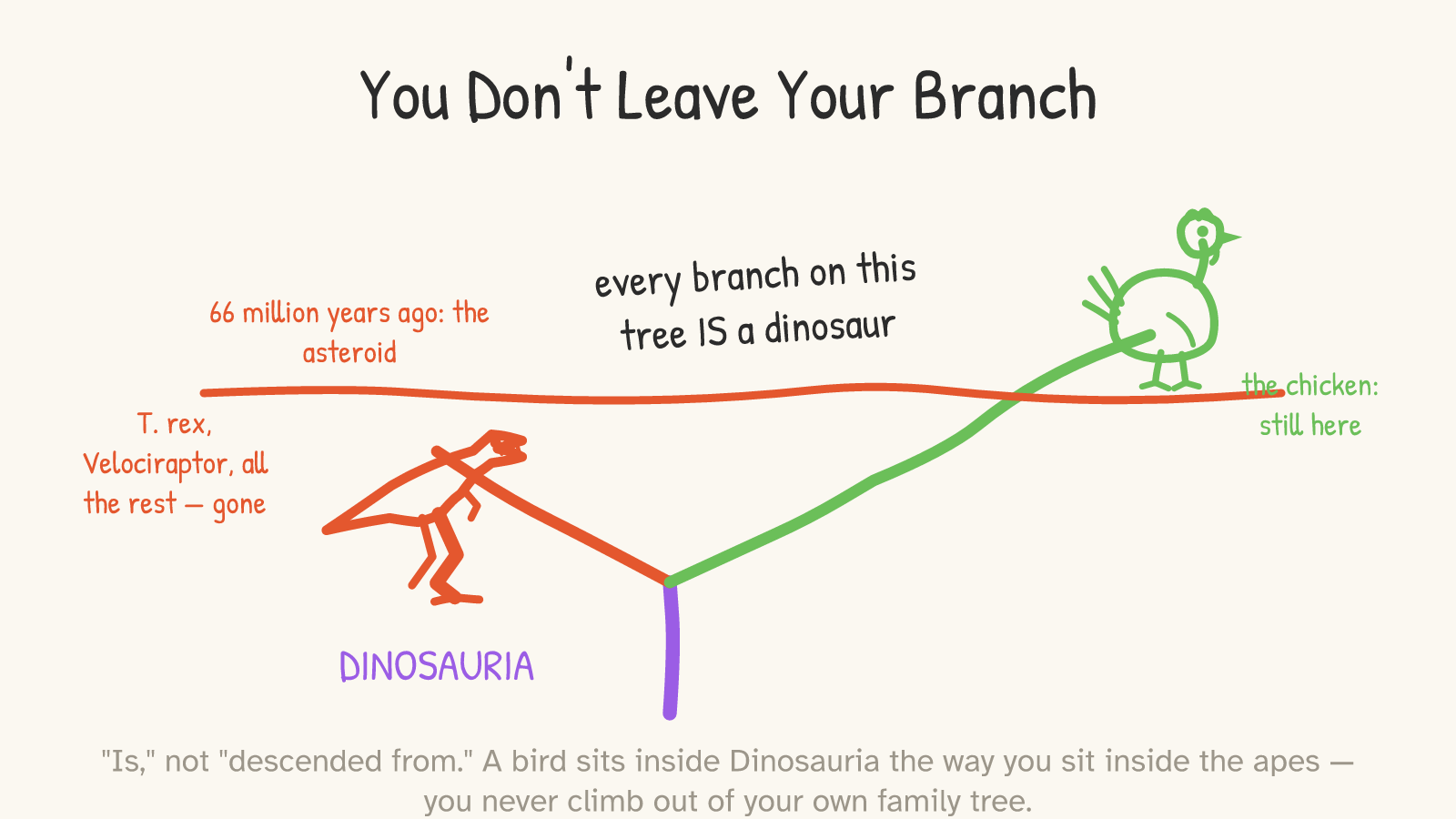
So when the dinosaur family tree branches and re-branches — Dinosauria splitting into the long-necked giants and the meat-eating theropods, the theropods narrowing down through the feathered maniraptorans to the first birds — every twig on that tree is still inside Dinosauria. Tyrannosaurus is on it. Velociraptor is on it. And the twig that kept going, past the catastrophe that killed the others, ends in a chicken. The chicken sits inside the dinosaurs the way you sit inside the apes: not next to them, not after them — within.2
A quick objection, cleared on the way past, because skeptics reach for it:
“Then why aren't crocodiles dinosaurs? They're reptilian, they're ancient, they're right there.”
Because crocodiles branch off just below the dinosaurs — they're the nearest living cousin, the outgroup. The dividing line of Dinosauria falls between them and everything descended from the first dinosaur, and that line is anatomical, not just a label: it's drawn at the specific hip and ankle architecture that gives dinosaurs their upright, legs-under-the-body stance. Birds inherited that architecture; crocodiles, with their sprawling gait, sit just outside it. It's the single most-tested branching point in the family tree, and birds land firmly on the dinosaur side of it.3
This isn't a recent fashion, either. The English anatomist Thomas Henry Huxley looked at a little dinosaur called Compsognathus and the first known bird, Archaeopteryx, back in the 1860s and concluded that birds were essentially modified reptiles — continuous with the dinosaur line.4 The idea then got buried for forty years over a single missing bone (we'll get to it), revived in 1969 when John Ostrom noticed how bird-like the raptor Deinonychus was, and finally nailed down in 1986 when Jacques Gauthier ran the whole question through the formal machinery of cladistics and birds came out, unambiguously, inside the theropods.5 The conclusion is old, it was resisted hard, and it kept winning.
Part 2It kept failing the tests
"Fine," says the fair-minded skeptic, "but a classification is just a decision about names. You've told me biologists draw the box around birds and dinosaurs together. Why did they draw it there?"
This is the right question, and the answer is bones. Not feathers — we'll get to feathers, and they're not even the strongest evidence. The skeleton alone does the work.
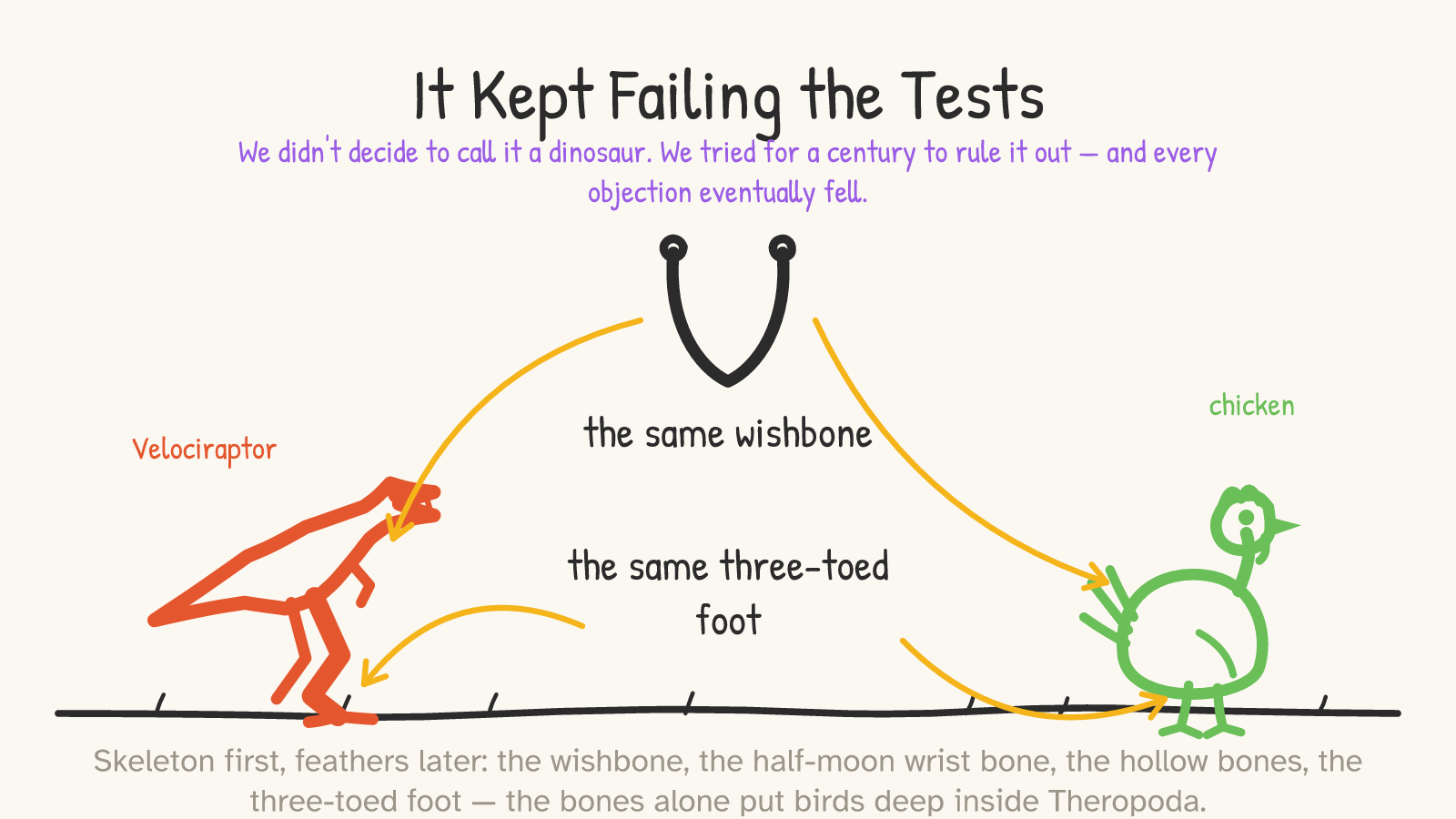
Pull a wishbone out of your roast chicken at dinner. That bone — the furcula, two collarbones fused into a springy V — is a theropod dinosaur trait. It's in Velociraptor. It's in Tyrannosaurus. It shows up in theropods as far back as the Triassic.6 (The forty-year burial of Huxley's idea happened because an influential 1926 book pointed out that no theropod clavicle was then known, and concluded birds must have come from something else. The bone was simply found later, over and over. The objection didn't survive contact with more digging.7) Then there's the half-moon-shaped wrist bone — the semilunate carpal — that lets a bird fold its wing; it's there in the raptors, doing the same folding motion that would later become the flight stroke.8 Hollow, air-filled bones plumbed by air sacs: theropod. A hinge-like ankle and a three-toed foot you'll recognize from any track in the mud or any chicken in any yard: dinosaurian.9
The point is not one lucky resemblance. When paleontologists build the big formal datasets that sort out who's related to whom, they score the whole skeleton across enormous character sets — one published matrix covers 853 anatomical features and 150 species — and birds settle deep inside the theropods no matter which subset of the data you use.10 That's the robustness that matters: you can throw out the feathers, throw out any single bone, throw out whole regions of the skeleton, and the answer doesn't move. The name follows the evidence. We didn't decide a chicken was a dinosaur; every attempt to find a reason it wasn't eventually collapsed.
Part 3The feather bonus
Now the feathers — because they're the part everyone's heard about, and they're genuinely spectacular, but I want to be careful about what they prove.
“Feathers don't make something a dinosaur. Lots of unrelated things converge on similar features. Bats and birds both fly; that doesn't make a bat a bird.”
Correct, which is exactly why Part 2 led with the skeleton and not the plumage. Feathers are corroboration, not the load-bearing wall. But what corroboration they are.

Starting in 1996, a series of fossils came out of the Yixian beds in northeastern China that settled an old argument in the most direct way possible: dinosaurs with feathers. Not birds — dinosaurs, with the full theropod skeleton, wearing feathers.11 Sinosauropteryx, a small predator, had a fuzzy coat of filaments. Caudipteryx had proper vaned feathers on stubby arms it could not possibly have flown with. Microraptor had flight-shaped feathers on all four limbs. And then, in 2012, the gut-punch: Yutyrannus, a tyrannosaur relative about nine meters long and well over a tonne, preserved with a shaggy coat of downy filaments.12 A feathered cousin of T. rex, far too big to fly, covered in the same kind of structure as the down on a chick.
Which tells you something the cartoons never did: feathers appear in the fossil record long before flight, on animals that couldn't have flown with them. Whatever they were originally for — insulation, display, something else still argued over — they were inherited by the fliers, not invented by them. So when you find the same feather on a ground-bound giant and on the chicken in the coop, the feather isn't a coincidence the chicken stumbled into. It's an heirloom — passed down the same branch, from dinosaurs that never left the ground to a bird that wears it still.
The behavior came down the branch too. We've found dinosaurs like Citipati fossilized sitting on their egg clutches in the exact splay-armed brooding posture of a nesting hen, killed and buried mid-incubation.13 And inside the leg bone of a female Tyrannosaurus, scientists found medullary bone — the special calcium-storing tissue that female birds lay down before they lay eggs. It was first known only in birds; it has since turned up in other dinosaurs too, exactly as you'd predict if the whole lineage shared the same egg-laying physiology.14 The chicken brooding her eggs is not imitating a dinosaur. She's doing what her lineage has always done.
One more strand, and this one I'm going to hand you with the caveats attached, because honesty about evidence is the whole game. In 2007, a team reported recovering fragments of collagen protein from a 68-million-year-old T. rex bone and found that, of all living animals, the sequence matched birds most closely — chickens and ostriches.15 It made headlines: molecules, not just bones, linking T. rex to chicken. But the result has been seriously contested. Critics argued the tiny peptide fragments could be contamination — from lab reagents, from bacteria, even from an ostrich sample processed in the same lab — and the debate is genuinely unresolved.16 So treat it as a suggestive footnote, not a pillar. The good news for the thesis is that it doesn't matter: the case was already overwhelming on bones alone. The molecules would be a nice bonus if they hold up. The skeleton doesn't need them.
Part 4But then I'm a fish
Here is the objection I promised you — the strong one, the one a sharp and scientifically literate person actually raises. It deserves the best version of itself:
“This is a cladistics parlor trick. 'Dinosaur' is a clade — a branch on a tree — and you're exploiting the gap between how biologists use the word and how everyone else does. By the same logic, I'm a lobe-finned fish, because tetrapods are nested inside the fish family tree. You can call a chicken a dinosaur and a human a fish and a whale a fish too, and every sentence is 'technically true' and completely useless for telling anyone what these animals are actually like. You've smuggled the menace of Jurassic Park onto a barnyard bird to manufacture wonder.”
This is good, and I'm going to concede the half of it that's right before I take it apart.

The half that's right: yes, "dinosaur" is a branch on a tree, and yes, by exactly this logic, you are a lobe-finned fish. Your lineage is nested inside the Sarcopterygii; you carry the body plan — the bones in your limbs map onto the bones in a fish's fin. If the whole claim were nothing but the naming convention, the skeptic would win, and we'd both go home.
But it isn't the naming convention. Look back at Parts 2 and 3. Birds were not assigned to the dinosaurs by fiat and then called dinosaurs as a quirk of vocabulary. They were placed there, against decades of resistance, by a mountain of physical evidence — the wishbone, the wrist, the foot, the deep skeletal anatomy, the feathered cousins in the ground. The name is the conclusion, not the premise. "You are a fish" works the same way and it isn't dishonest either — it's just unfamiliar, and unfamiliar is not the same as false. You really do carry the fish's architecture. The chicken really does carry the dinosaur's.
So why prefer the cladistic word "dinosaur" — the one that includes the chicken — over the everyday one that means "the big extinct monsters"? Because the cladistic group tracks something real: a specific cluster of inherited traits that the chicken demonstrably has. The everyday version, "dinosaurs are everything in the group except birds," is the one that has to do something arbitrary — it cuts a single natural family in two, keeping the extinct forms while discarding the living descendants that have the very same skeleton, with the cut falling wherever feels intuitive rather than at any real boundary in the biology. That's the move that puts the naming ahead of the biology. Pointing at the inheritance and naming it accurately is the opposite of a trick.
I'll concede one honest thing to the skeptic, though: the jolt in "you're eating a dinosaur" borrows some of its voltage from the word's pop-culture baggage — the T. rex, the roar, the movie. Strip that away and the literal content is quieter and truer: this animal carries a specific ancient anatomy, and its lineage walked through an extinction that erased almost everything else. The label is the doorway. The anatomy and the survival are the actual story — and they're enough.
And this is the place to bury the last serious scientific resistance, because there was some, and the skeptic deserves to know it lost on the merits, not by decree. A camp of researchers — the "birds are not dinosaurs" school, associated with ornithologists like Alan Feduccia and Larry Martin — held out for decades, and they raised several real anatomical objections. The most technically pointed was the fingers.
“Bird embryos look like they grow fingers two-three-four, but theropod dinosaurs had fingers one-two-three. If the hands are built from different fingers, the bird hand can't be the dinosaur hand, and the whole thing collapses.”
For a while that was a real problem. It was addressed in 1999 by the "frameshift" hypothesis — the idea that the developmental program building the fingers had shifted over by one position, so what develops in the two-three-four spots grows into the identities of fingers one-two-three — and later work on which genes switch on in the developing digits is consistent with that solution, though the exact developmental mechanism is still being worked out.17 The other big objection, that Archaeopteryx was older than the raptors it supposedly descended from (the "temporal paradox"), dissolved when feathered dinosaurs older than Archaeopteryx — like the ~160-million-year-old Anchiornis — came out of the ground, showing the bird-line branches of the theropod tree are themselves older than Archaeopteryx.18 The holdouts didn't lose because the field stopped listening. They lost because every objection they raised got a specific, evidenced answer, and no positive evidence for any alternative ancestor ever showed up.
Part 5Not a shrunken T. rex
Before this tips into hype, let me draw the lines the thesis does not cross, loudly, because the overreach is real and a careful reader should hold me to the modest claim.
“So you're telling me my dinner is a tiny T. rex.”
No. And anyone who tells you that is overselling it.

A chicken is not descended from Tyrannosaurus. T. rex is a cousin, not a grandparent. Their two lineages — the tyrannosaurs and the bird-line — split apart in the Middle Jurassic, on the order of 165 million years ago (the estimates run from about 155 to 175), and then evolved independently for longer than the entire age of the mammals.19 Calling a chicken a "shrunken T. rex" is exactly as wrong as calling you a "shrunken baboon": you and the baboon share an ancestor, you're both primates, but you didn't descend from one and neither did your dinner from T. rex. The honest sentence is "a chicken is a living dinosaur, in the same broad group as T. rex." That's precise. "Domesticated T. rex" is a T-shirt slogan, and I'm disowning it.
And there's a second limit, which is really the heart of the wonder rather than a caveat against it. Almost no dinosaurs made it. Sixty-six million years ago an asteroid struck, the world burned and went dark, and every non-bird dinosaur died — along with most of the birds. The great diversity of Mesozoic birds, the toothed ones and the clawed ones, went extinct right alongside T. rex. Only a few beaked, ground-dwelling, seed-eating birds squeaked through the bottleneck, and every bird alive today — every sparrow, every eagle, every chicken — descends from that handful of survivors.20 So "chickens are dinosaurs" really means "chickens are the surviving dinosaurs," the far tip of the one twig that didn't burn. That's not a deflation of the claim. That's the most astonishing part of it.
Part 6We tamed the survivor
Which brings us to the part that is genuinely our doing. The dinosaur survived the asteroid on its own. The part where it ended up in a coop — that was us.
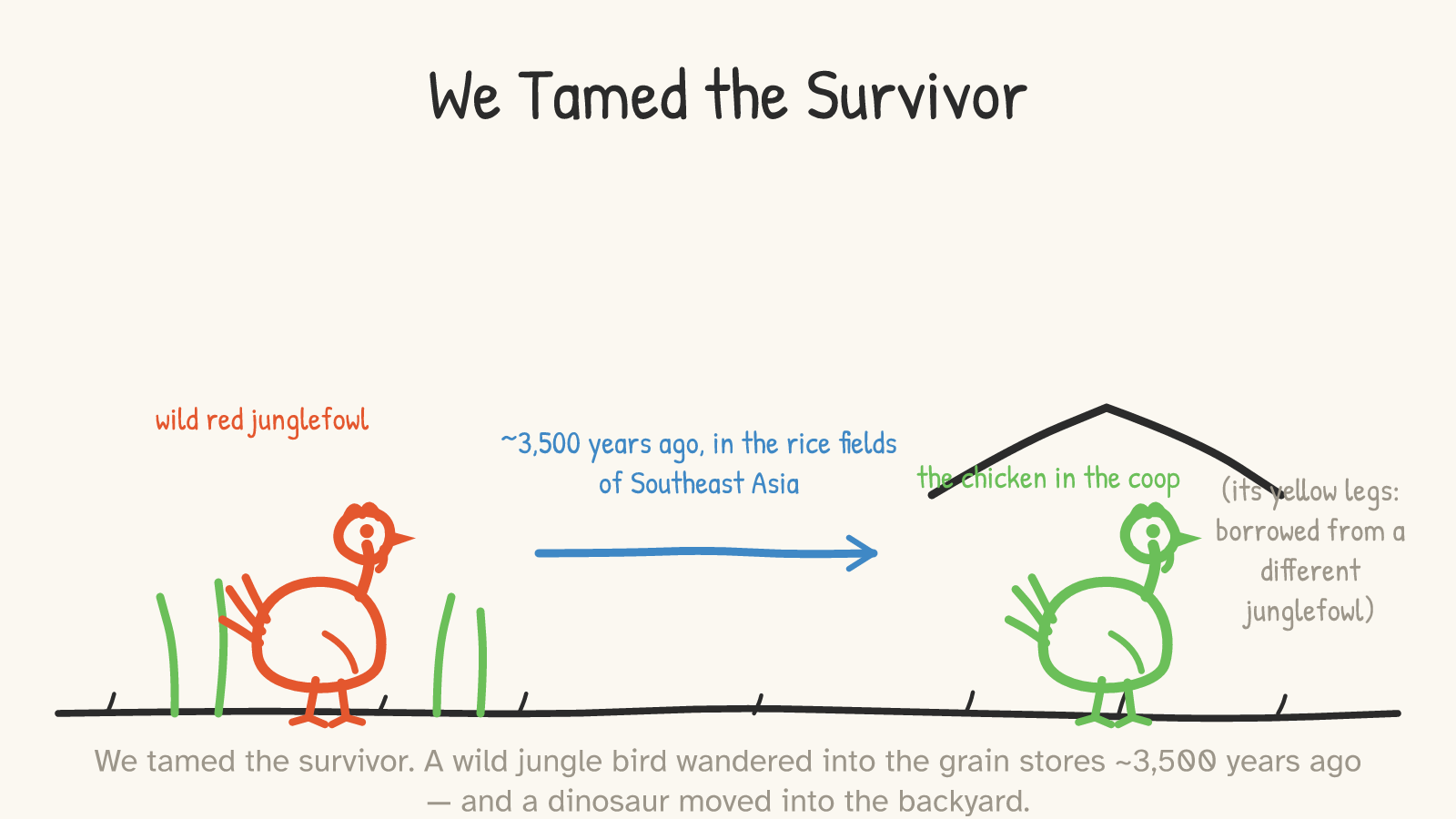
The chicken's wild ancestor is the red junglefowl, a gorgeous, wary, tree-roosting bird of the South and Southeast Asian forests, about the size of a slim bantam.21 The domestication story has actually been rewritten in the last few years, and the honest version is more interesting than the old one. For a long time textbooks said chickens were domesticated eight or ten thousand years ago in northern China or the Indus Valley. A careful reassessment in 2022 threw most of that out: many of the "early" bones turned out to be pheasants, or wild birds, or simply undated. The revised, better-supported story is that domestication began only about 3,500 years ago, in mainland Southeast Asia — at sites like Ban Non Wat in central Thailand — when the spread of dry-rice and millet farming drew wild junglefowl in to feed on the grain, and a wary forest bird gradually became a tame one living alongside people.22 It's a young relationship. The dinosaur is 165 million years old; the domestic part is younger than the pyramids' middle age.
A small, lovely detail that shows how much tinkering we did: your chicken's yellow legs and yellow skin aren't from the red junglefowl at all. That trait was borrowed — introgressed, the geneticists say — from a different species, the grey junglefowl, somewhere along the way.23 And the tinkering accelerated to a sprint in living memory. Raise a 1950s broiler chicken and a modern one side by side on the same feed, and the modern bird grows to roughly four to five times the weight at the same age — a body reshaped more in seventy years than in the previous three thousand.24 We took a surviving dinosaur and, very recently and very deliberately, turned it into the most efficient meat-growing machine the lineage has ever produced.
Part 7The most numerous dinosaur that ever lived
Stand back and look at what that produced, because the scale is the punchline.

There are about 33 billion chickens alive on Earth at any given moment — a number that has roughly tripled since 1990.25 That makes the chicken, by a margin that isn't close, the most numerous bird on the planet; the nearest wild competitor numbers perhaps 1.5 billion.26 We raise and slaughter more than 70 billion of them a year.27 By sheer biomass and headcount, the domestic chicken is very likely the most numerous dinosaur that has ever existed in the 165-million-year history of the group — and certainly the most numerous bird.
This is so true that geologists have started to take it literally. A 2018 paper argues that the broiler chicken — its bones reshaped by breeding, scattered in landfills across every continent in numbers nothing else comes close to — will be one of the defining fossil signatures of the human epoch in the rock record. A future geologist, reading the strata, will find a thin global layer marking our time, and it will be made of dinosaur bones: ours, the ones we made.28
So here is where the wonder finally settles, and it isn't in a museum or a movie. It's in the most ordinary place there is. The dinosaurs did not all die. One kind ducked under the catastrophe, rode out the dark, diversified into every sky on Earth — and then, 3,500 years ago, a few of them followed the grain into our villages and never left. They are standing in backyards right now. They are roosting by the billions. One of them is on a shelf in your refrigerator.
When you eat chicken, you are eating a dinosaur — the most numerous bird, and very likely the most numerous dinosaur, that has ever lived, though only because of what we did to it. The whole time, the most extraordinary fact about the animal on your plate was hiding in the blandest corner of your kitchen.
Footnotes & receipts
- Monophyly / the clade principle: a clade is an ancestor plus all its descendants, so a lineage cannot evolve "out of" its ancestral clade — bats remain mammals, whales remain artiodactyls. Standard in any systematics text (e.g., Padian & Chiappe, Scientific American / review literature, 1998). Tier 1/2. ↩
- Phylogenetic placement: Dinosauria → Saurischia → Theropoda → Coelurosauria → Maniraptora → Avialae → Aves; the domestic chicken Gallus gallus domesticus sits within crown birds (Neornithes), hence is a maniraptoran theropod. Consensus review: Brusatte et al., "The origin and diversification of birds," Current Biology 25 (2015). Tier 1. ↩
- Crocodylians are the extant outgroup to dinosaurs within Archosauria (the bird-line and croc-line split below Dinosauria); birds fall inside Dinosauria, crocs outside. Standard archosaur phylogeny (Nesbitt 2011, Bull. AMNH). Tier 1. ↩
- T. H. Huxley, 1860s work arguing the bird–reptile/dinosaur link from Compsognathus and Archaeopteryx; his 1868 paper "On the animals which are most nearly intermediate between birds and reptiles," Geological Magazine, is the canonical statement. His position is paraphrased here, not quoted: the widely circulated wordings ("an extremely modified and aberrant Reptilian type," 1863; "greatly modified Reptiles," attributed to an 1867 address) appear in secondary sources (Clark University Huxley File; Linda Hall Library "Paper Dinosaurs") but were not traced to a primary scan, so no direct quotation is asserted. Tier 2. ↩
- History of the idea: G. Heilmann, The Origin of Birds (1926, thecodont hypothesis); J. Ostrom on Deinonychus (1969, Bull. Peabody Mus.) reviving the theropod link; J. Gauthier, "Saurischian monophyly and the origin of birds," Mem. Calif. Acad. Sci. (1986), the foundational cladistic analysis placing Aves within Coelurosauria. Tier 1/2. ↩
- Furcula (wishbone) in theropods including Triassic forms; distribution documented in Nesbitt et al. (2009) and earlier. The chicken's wishbone is the homolog of the theropod furcula. Tier 1. ↩
- Heilmann's 1926 clavicle objection: no theropod furcula was known to him, leading him (wrongly) to reject theropod ancestry; subsequent finds of theropod furculae nullified the objection. Tier 1/2 (history of science). ↩
- Semilunate carpal (half-moon wrist bone) in maniraptoran theropods, enabling the folding wrist motion that is the mechanical precursor of the avian flight stroke; described by Ostrom and Gauthier; see also Xu et al. on wrist evolution (2014, Scientific Reports / related). Tier 1. ↩
- Pneumatized (air-filled) bones with air sacs, mesotarsal hinge ankle, and tridactyl (three-functional-toed) foot as theropod/dinosaurian features retained in birds. Standard osteology; The Dinosauria, 2nd ed. (Weishampel et al., 2004). Tier 2. ↩
- Large cladistic matrices: e.g., Turner, Makovicky & Norell, "A review of dromaeosaurid systematics and paravian phylogeny," Bull. AMNH 371 (2012), scoring 853 characters across 150 taxa. The "hundreds of shared characters" claim refers to total matrix size and the robustness of the bird-within-theropod result across analyses, not to hundreds of features each uniquely diagnostic of the link. Tier 1. ↩
- Feathered non-avian dinosaurs, Yixian/Jehol Group, China: Sinosauropteryx (Ji & Ji 1996, first filamentous integument on a non-avian dinosaur); Caudipteryx (Ji et al. 1998, Nature, pennaceous feathers, flightless oviraptorosaur); Microraptor (Xu et al. 2003, Nature, four feathered limbs). All Nature-class. Tier 1. ↩
- Yutyrannus huali, Xu et al. (2012, Nature 484:92): ~9 m, ~1.4-tonne tyrannosauroid with filamentous feathers — feathers on the tyrannosaur lineage, on an animal far too large to fly. Anchiornis (Hu et al. 2009) and others extend feathers across coelurosaur groups. Tier 1. ↩
- Brooding posture: Citipati osmolskae ("Oviraptor") specimens preserved atop egg clutches in an avian brooding posture, Norell et al. (1995, Nature 378:774). Tier 1. ↩
- Medullary bone (the calcium-mobilizing tissue female birds form before laying) identified in a Tyrannosaurus rex femur: Schweitzer et al. (2005, Science 308:1456); chemically corroborated in Schweitzer et al. (2016, Scientific Reports 6:23099). Originally known only in living birds, it has since been reported in other non-avian dinosaurs — including the theropod Allosaurus and the ornithopod Tenontosaurus (Lee & Werning, 2008, PNAS 105:582) — consistent with a shared dinosaurian egg-laying physiology, and more widely accepted than the collagen result in note 15. Tier 1. ↩
- T. rex collagen peptides: Asara et al. (2007, Science 316:280) recovered short collagen sequences from a ~68-Myr T. rex (MOR 1125); phylogenetic analysis grouped it closest to birds (chicken, ostrich) among living taxa; Schweitzer et al. (2009, PLoS ONE) reported a similar result for an 80-Myr hadrosaur. Tier 1 — but see note 16. ↩
- Contamination critique: Buckley et al. (2008) and Pevzner et al. (2008, J. Proteome Res.) argued the peptides could be contamination (lab reagents, bacterial biofilm, or an ostrich sample processed in the same facility); a reanalysis found several T. rex sequences also present in ostrich collagen. The reliability of Cretaceous-age protein sequencing remains contested as of the 2020s. This essay treats the molecular result as a contested corroborating strand, not as primary evidence; the morphological case stands without it. Tier 1. ↩
- Digit homology / the "frameshift" hypothesis: Wagner & Gauthier, "1,2,3 = 2,3,4: A solution to the problem of the homology of the digits in the avian hand," PNAS 96 (1999); supported by later developmental-genetic work on digit-identity gene expression (e.g., Vargas & Fallon, 2005). Resolves the apparent II–III–IV vs I–II–III mismatch in favor of homology with the dinosaur hand. Tier 1. ↩
- Temporal paradox: the apparent problem that Archaeopteryx (~150 Ma) predated its supposed dromaeosaur relatives dissolved with the discovery of Jurassic paravians as old or older — Anchiornis huxleyi, ~160 Ma (Hu et al. 2009, Nature); statistical treatment of sampling gaps in Brochu & Norell (2000). Tier 1. ↩
- Tyrannosauroid–bird-line divergence within Coelurosauria estimated to the Middle Jurassic, ~165 Ma (analyses range ~155–175 Ma); see Holtz in The Dinosauria 2nd ed. (2004) and molecular/morphological timetrees. Chicken and T. rex share a theropod ancestor but neither descends from the other. Tier 1/2. ↩
- K–Pg survival: all non-avian dinosaurs and most Mesozoic bird lineages (enantiornithines, hesperornithines, etc.) went extinct at 66 Ma; only crown birds (Neornithes) survived. Longrich et al. (2011, PNAS 108:15253) on the mass extinction of birds at the boundary; Field et al. (2018, Current Biology) on the ground-dwelling, beaked, generalist ecology of the survivors. All living birds descend from that bottleneck. Tier 1. ↩
- Wild ancestor: red junglefowl Gallus gallus; domestic ancestry overwhelmingly G. gallus per whole-genome studies (e.g., Wang et al. 2020, Cell Research 30:813). Tier 1. ↩
- Domestication date and place (revised), two companion 2022 papers from the same research group: J. Peters et al. (first author Joris Peters), "The biocultural origins and dispersal of domestic chickens," PNAS 119:24 (2022) — the main large-scale reassessment (remains from 600+ sites in 89 countries) placing unambiguous domestic chickens no earlier than ~1650–1250 BCE (~3,500 years ago) in central Thailand / mainland Southeast Asia, tied to dry-rice/millet cultivation, with many earlier "chicken" bones reidentified or undated; and J. Best et al., "Redefining the timing and circumstances of the chicken's introduction to Europe and north-west Africa," Antiquity 96 (2022), the companion redating of the European dispersal. The older "8,000–10,000 years" figure is superseded. Genomic bottleneck estimates (~3,000–4,000 yr) agree. Tier 1. ↩
- Yellow-skin allele introgressed from the grey junglefowl Gallus sonneratii: Eriksson et al. (2008, PLoS Genetics 4:e1000010). The only well-documented cross-species genetic contribution to the domestic chicken. Tier 1. ↩
- Broiler transformation: Zuidhof et al. (2014, Poultry Science 93:2970) raised 1957, 1978, and 2005 commercial broiler lines under identical conditions; the 2005 line reached roughly four to five times the body mass of the 1957 line at the same age (~0.9 kg vs ~4.2 kg at 56 days), with growth rate up ~400%. Applies to commercial broilers specifically, not heritage or laying breeds. Tier 1. ↩
- ~33 billion chickens alive at any time: FAO / FAOSTAT data (global stock ~33 billion in 2020, up from ~10.6 billion in 1990); compiled e.g. by the Carnegie Museum of Natural History (citing FAO). "Alive at any moment" is a stock figure. Tier 2/3. ↩
- Most numerous bird by orders of magnitude; nearest wild bird, the red-billed quelea, estimated ~1.5 billion. Tier 3. ↩
- >70 billion chickens slaughtered per year: FAO (≈73–77 billion in recent years; higher if male-chick culling is included). Annual slaughter exceeds the live stock because broilers turn over in ~6 weeks. Tier 2. ↩
- Broiler chickens as an Anthropocene marker: Bennett et al., "The broiler chicken as a signal of a human reconfigured biosphere," Royal Society Open Science 5:180325 (2018). Argues broiler bones — morphologically distinct, globally distributed, hugely abundant — will form a defining stratigraphic signature of the present epoch. Tier 1. ↩